Dehydroepiandrosterone, Melatonin, and
Testosterone in Human Evolution
"Androgens in Human Evolution" is derived from this article.
Copyright ã 1995, 1996. Revised from 1985 by James Michael Howard.
This thesis is an entirely new explanation of
human (hominid) evolution and part of a new paradigm. I believe hominids
evolved because of two primary changes: (1) increased production of testosterone
in both sexes and (2) increased use by the brain of the pineal gland hormone,
melatonin, and the adrenal gland hormones, dehydroepiandrosterone (DHEA) and
the more abundant form, DHEA-sulfate (DHEAS). This explanation of human
phylogeny also provides some new explanations of human ontogeny, pathology, and
behavior. These ideas will be presented in a series of articles.
Relatively recent hominid fossil finds
indicate that bipedal locomotion occurred early in human evolution and may have
developed in an arboreal environment (National Geographic, Sept., 1995,
page 38.) Bipedal walking preceded large increases in brain size by millions of
years and occurred near the separation of chimpanzee and human lines. I suggest
the same mechanism that produced bipedal walking eventually triggered the large
increases in brain size of later hominids.
My foundation hypothesis is that DHEA is
necessary for duplication and transcription of DNA. Therefore, all growth,
development, maintenance, activation, and aging are dependent upon production,
antagonism, or loss of production of DHEA. All tissues compete for available
DHEA by "capturing" it from blood. (Serum, the liquid part of blood,
is used for optimum cell duplication and gene transcription in vitro.
DHEA is the major hormone found in serum.) The following chart represents the
availability of DHEA during the human life-span. (It is derived from a
combination of data in Adrenal Androgens, A.R. Genazzani, Raven Press,
1980.)
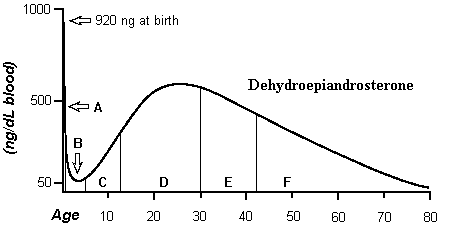
Period A is the first year; B is
from one year to adrenarche; C is childhood; D is the reproductive period; E
and F are redundancy, with E being reproductive redundancy and F, fatal
redundancy.
DHEA and the Brain
Metabolism
In the competition for DHEA among tissues,
nervous tissues capture DHEA better than other tissues. "Brain tissue
naturally contains 6.5 times more DHEA than is found in other tissues." (Total
Health, Feb. 1994, page 42, E.R. Braverman, M.D.) This is why brains
evolved; nervous tissues grow at a greater rate than other tissues. For
example, in a chicken embryo, after blood vessels form a system for nutrient
delivery, the brain is the first organ to develop. Since DHEA may be necessary
for transcription and the brain captures more DHEA than other tissues, the
brain should have the highest transcription rate. Additionally, DHEA should
affect all cellular DNA, hence, it should affect mitochondrial DNA, as well as
nuclear DNA. Mitochondria are the seat of metabolic activity in cells. The
following quotations support my hypotheses. In the second quotation,
"protein synthesis" indicates transcription has occurred.
"Brain is characterized by high
metabolic activity and exhibits two to three times the transcriptional activity
of other tissues." (Journal of Neurochemistry 1991; 56: 812)
"These findings indicate that
mitochondrial respiration is the earliest factor affected by DHEA and may be
associated with protein synthesis." (Journal of Nutrition 1991; 121:
240)
Brain Growth and
Puberty
The brain's increased ability to capture DHEA
results in its growth at the expense of other organs. This explains why:
"...the brain is most unusual in its pattern of accelerated growth in
comparison to the other organs and to the body as a whole." (Patterns
of Human Growth, B. Bogin, Cambridge University Press, 1988, page 61.) The
brain's use of DHEA produces Period B of the chart above. Period B is a decline
in measurable DHEA, i.e., the brain uses so much DHEA for growth and
development that DHEA levels decline. At birth, DHEA is produced in extremely
large amounts for early brain, and body, growth; this is Period A. (In children
who succumb to SIDS, it is found that "Somatic [body] growth and brain
weight were significantly greater in SIDS than controls." (Journal of
Neuropathology and Experimental Neurology 1991; 50: 29.) This would
reduce DHEA of Period B so low that enough is not available to maintain
activation of the brainstem which controls the heart and breathing.) As the
brain approaches final development, DHEA levels increase. This is the beginning
of Period C; it is called adrenarche. Puberty occurs at the end of Period C.
Earlier, I mentioned that DHEA is used for "activation." As the brain
finishes growth and development, it starts using the extra DHEA for higher
functional activity (thinking, behavior, etc.). The most pronounced behavior
following adrenarche and childhood is puberty. I suggest puberty simply occurs
when use of available DHEA switches from growth and development to reproductive
drives.
Pathology
In vitro, extremely small amounts of DHEA increase neuron
differentiation and survival (Journal of Neuroscience Research 1987; 17:
225.) The neuron is the basic building block of the brain. During development in
utero or postnatally, low levels of, or antagonism of, DHEA availability
should adversely affect brain growth. This should occur along a continuum,
i.e., in severe cases this could lead to anencephaly (lack of brain
development) to less subtle forms of brain development that are exposed only by
reduced DHEA availability later in life. For sake of this section, I will
briefly demonstrate the connection of DHEA with two of these that cause a lot
of problems and costs in our society: schizophrenia and Alzheimer's disease. (I
will consider these in greater detail later in this series.)
In 1985, I proposed that DHEA should be low
in Alzheimer's (A Theory of the Control of the Ontogeny and Phylogeny of Homo
sapiens by the Interaction of Dehydroepiandrosterone and the Amygdala,
Copyright, 1985, James Michael Howard.) This was supported in The Lancet;
1989, Sept. 2, page 570 and in Biological Psychiatry 1991; 30:
688, as well as other journals. The point is that DHEA naturally begins to
decline in Period E. This is the time when early Alzheimer's disease occurs, in
people genetically predisposed to neuropathy during this drop in availability
of DHEA. If I am correct, dementia of this type should occur in the normal
population as DHEA declines in late redundancy (Period F). Depending on the
individual, one might develop some form of senile dementia during old age.
Therefore, it should be, and is, a common form of dementia in the normal, very
old, population.
"Of those over the age of 65
years, an estimated 10.3% had probable Alzheimer's disease [AD]. The prevalence
rate was strongly associated with age. Of those 65 to 74 years old, 3.0%
[three] had probable AD, compared with 18.7% of those 75 to 84 years old and
47.2% of those over 85 years. Other dementing conditions were uncommon.
...These data suggest that clinically diagnosed AD is a common
condition..." (Journal of the American Medical Association 1989; 262:
2551)
I also propose that schizophrenia develops as
a consequence of low DHEA. Schizophrenia exhibits significantly reduced levels
of DHEA (Biological Psychiatry 1973; 6: 23), and "the
schizophrenic group was found to have significantly less gray matter than the
control group ...in all six cortical subregions analyzed..." (Archives
of General Psychiatry 1992; 49: 195.) (Neurons compose the majority
of "gray matter.") Reduced DHEA reduces brain growth. Schizophrenia
represents a disorder of reduced DHEA and reduced development, later exposed by
antagonism of DHEA availability by the hormones, testosterone and cortisol. The
point of this data is that DHEA is intimately connected to brain growth and
function, and hominid brain evolution can be tied to increases in DHEA
availability.
DHEA in a Monkey, the Chimp, and Humans
Use of DHEA by the brains of monkeys,
chimpanzees, and humans generate differences in the life-span charts of DHEA
that follow a pattern. Use of DHEA by the large brains of humans causes the
lowest measurable levels of DHEA of Period B. Period D in humans is lower
because of use of DHEA for brain function; larger brains use more. Therefore,
monkeys have the least Period B and lowest Period D. Chimpanzees are between
the two, but nearer to humans; the chimp is the only animal which exhibits an
adrenarche similar to humans (Endocrinology 1978; 103: 2112.)
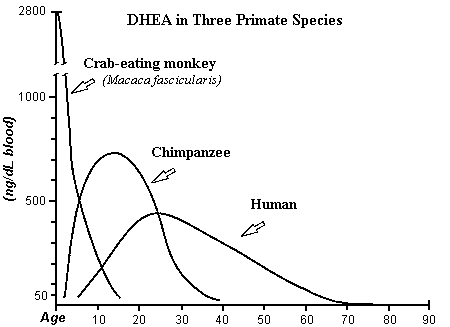
This figure is derived from the
first chart (human DHEA); Journal of Reproduction and Fertility 1985; 74:
347, from Text- fig. 5, page 355 (monkey); and J. Repro. Fert. Supplement
No. 28; 1980, from Text-fig. 5, page 137 (chimpanzee)
Period B occurs in the monkey, but it is so
small and rapid in the scale of this chart that it cannot be seen. Since the
brain reaches final growth early in the monkey, puberty is reached early.
Evolution of the Hominids
Brain Size
The hominids follow a trend in development
that differentiates them from chimpanzee and gorilla lines (pongids). That is,
the ratio of brain size to body weight increases more in hominid evolution.
This is clear in the following chart adapted from Human Evolution An
Illustrated Introduction, R. Lewin, W.H. Freeman and Company, New York,
1984; after figure on page 81.
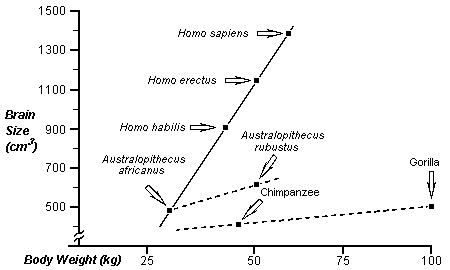
The trend of both hominids and pongids in
body weight is toward increased mass. Testosterone (T) increases body mass.
This suggests a shared selection pressure toward increasing T in both groups.
Testosterone is extremely important to human evolution, and human males and
females produce more testosterone than chimpanzee males and females,
respectively (J. Repro. Fert. Supplement No. 28, The Great Apes of Africa,
1980, see Text-fig. 2 (males), page 134 and Text-fig. 5 (females), page 137).
In hominids and pongids, increased testosterone produces a reproductive
advantage in male access to females. Testosterone produces massiveness and
aggressiveness.
Testosterone will increase in a group,
because it increases sexual opportunity. Aggressive, high T males force less
aggressive, less massive males away from females. Over time, therefore, T
increases in hominids and pongids; this is the driving force that causes all
lines in the chart above to move to the right, i.e., increased size. This is
supported in the fossil record; early Australopithecines were smaller:
"...it is notable that the more ancient Australopithecines had thin skull
bones and only modest protuberances on his cranium" (Encyclopædia
Britannica 1984; 8: 1033.) Eventually, extremely massive males, such
as Australopithecus robustus and A. boisei, were produced. These
species became extinct without contributing to the hominid line; too much
testosterone is a bad thing.
The "pushing" of lower testosterone
males away from the group can be seen in living primates. Here is an example
from Wickings E.J. et al., "Testicular Function, Secondary Sexual
Development, and Social Status in Male Mandrills (Mandrillus sphinx)" Physiology
& Behavior 1992; 52: 909.
"Positive correlations between
dominance rank and plasma testosterone levels have been described for adult
males of several primate species in captivity, but the relevance of such
observations to free-ranging animals is unclear. CIRMF in Gabon maintains a
breeding group of 45 mandrills in a six hectare, naturally rainforested
enclosure. This study describes correlations between dominance rank (in
agonistic encounters), levels of plasma testosterone, testicular volume, body
weight, and development of secondary sexual characteristics (red and blue
sexual skin on the muzzle and rump areas) in male mandrills under semifree
ranging conditions. Two morphological and social variants of adult male
mandrill were identified. Large-rumped or fatted adult males (n = 3) remained
in the social group and exhibited maximal development of sexual skin coloration
as well as large testicular size and highest plasma testosterone levels. By
contrast, slimmer-rumped or nonfatted males (n = 3) lived a peripheral or
solitary existence and these exhibited less development of their secondary
sexual coloration and had smaller testes and lower plasma testosterone levels.
Longitudinal studies of gonadal development in these six males revealed that
testicular volumes and plasma testosterone levels increased most rapidly during
pubertal development (4-5 years of age) in the three animals which proceeded to
the fatted condition. These included the highest ranking, group-associated male
which exhibited the most intense sexual skin coloration and had higher
testosterone levels, although this was not correlated with testicular volume.
This study shows that in the male mandrill social factors and reproductive
development are interrelated."
All lines in the chart above also move
upward, i.e., all groups show an increase in brain size. This is also partly
due to increasing testosterone. I suggest T acts by increasing the uptake of
DHEA in testosterone-target tissues. That is, T increases the uptake of DHEA
for transcription of T-activated genes. The brain is full of T receptors used
for capturing T. Therefore, increases in T will increase the supply of DHEA to
the brain. T receptors are located in the cerebral cortex, but mainly in
subcortical regions of the brain. This means that increases in T will only
increase brain size by a limited amount over time. However, this effect of T
does occur in humans. Males are exposed to T in utero and for a few
months postnatally. The result is a slightly increased head circumference in
males at birth and at the end of the first year (Sexual Dimorphism in Homo
sapiens A Question of Size, R.L. Hall, Praeger Publishers, New York,
1982, page 281.) However, significant increases in hominid brain size depend on
another mechanism of increasing the availability of DHEA.
DHEA increases resting metabolism; more heat
from dietary intake. Therefore, increased DHEA allows migration into colder
environments.
"There is growing clinical and
experimental evidence that dehydroepiandrosterone ...plays an important
regulatory role in intermediary metabolism by inhibiting the storage of dietary
energy as fat. For instance, one of the predominant features associated with a
DHEA deficiency in humans is obesity. ...Recently it has been reported that
these inhibitory effects of DHEA on adiposity can be attributed to an increase
in resting metabolism." (Journal of Nutrition 1987; 117:
1287)
Migrating groups of low testosterone would
have an advantage in colder climates only if they produced increased DHEA. That
is, they could produce more heat from scarce calories. Increased DHEA increases
brain growth and development, hence, groups of higher DHEA forced northward
should, on average, exhibit increased cranial size. In Asia, northern groups of
Homo erectus have larger brains than southern groups.
"Some difference in estimated
brain size is apparent between the Javanese and the Chou-K'ou-tien (Peking
Chinese) populations of Homo erectus. Thus, for seven Javanese crania,
the average is 833 cc, with a range from 750 to 1,030 cc; while for five
Chou-K'ou-tien crania, the capacity ranges from 915 to 1,225 cc and averages
about 1,043 cc. That is, the mean capacity in the Peking fossils of H.
erectus exceeds that of the Javanese by about 160 cc." (Encyclopædia
Britannica 1984; 8: 1032)
DHEA and Melatonin
I propose melatonin is directly involved in
DHEA production. This may be the mechanism of significant increases in brain
size found in northern hominids. These two hormones are directly linked to each
other in the sleep- wake cycle; one affects production of the other during this
cycle. DHEA is used during the day to activate consciousness and is literally
"used up." We get tired at the end of the day. This loss of DHEA
stimulation allows the pineal gland to release melatonin, synthesized earlier.
This large release of melatonin starts the first slow wave sleep of the night.
Melatonin triggers this sleep by slowing release of prolactin, which is known
to specifically stimulate DHEAS. During the night, melatonin is also used up;
then a large release of prolactin triggers the large morning release of DHEAS
that triggers awakening. I suggest this cycle is necessary for growth. (In the
case of SIDS, it may be that these children produce too much melatonin. This
would reduce DHEA to dangerously low levels during sleep. Melatonin is also low
in schizophrenia.)
Sunlight directly affects melatonin
production, i.e., decreased sunlight increases melatonin. Migration of hominids
northward increases melatonin and its effect on growth. (Migration of hominids
southward from the equator would produce the same effect.) I suggest melatonin
is directly involved with DHEA in brain growth. The following chart
demonstrates the connection of melatonin and DHEA. The time of greatest
melatonin production is also the time of greatest use of DHEA for brain growth
(Period B).
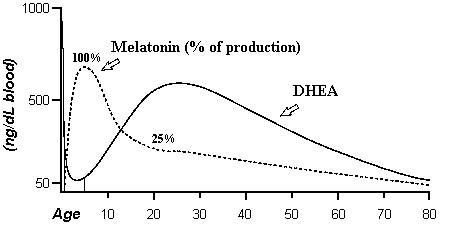
Bipedal Locomotion
and Other Changes
I suggest increasing testosterone levels
began a series of changes that resulted in bipedal walking. The mechanism
involves redirection of DHEA for use by T-target tissues at the expense of
other tissues. That is, as T increased, anatomical structures were merely
"remodeled" by increases in DHEA for T-target tissues, while other
tissues changed due to decreasing availability of DHEA. No mutations would be
required for this effect to occur. Many differences between males and females
occur as a result of differences in testosterone. Increases in T during
millions of years could produce differences in the fossil record.
Bipedal locomotion occurred millions of years
prior to any significant increases in brain size. However, in the early hominid
line, Australopithecus, some increase in brain size is found. This is
the testosterone effect on brain size, mentioned above. Along with this small
brain increase are changes which I attribute to remodeling caused by T. When
compared to pongids, Australopithecine canine teeth are nonprojecting and
reduced in size, their foramen magnum opens downward, and they are bipedal. In
Australopithecines, changes in the size and structure of the brain, induced by
testosterone in utero, could change the angle of the foramen magnum due
to plasticity in the developing skull. Taken together, these produce bipedal
walking and a shift away from teeth as weapons to hands as weapons of aggression.
These anatomical and functional changes are a consequence of increasing
testosterone.
Teeth are sensitive to DHEA availability.
Nature does not often reduce large, projecting canine teeth, very well adapted
as offensive weapons. I suggest reductions in size, or projection, of teeth
result from reduced DHEA availability. This reduction in teeth size directly
parallels increases in brain capacity. Chimpanzees have smaller brains,
therefore, they produce more available DHEA, and they have much larger,
projecting teeth. Reduced teeth size is a consequence of competition between
teeth and brain.
The possible connection of teeth and use of
DHEA by the brain is clearer in modern humans. There are two times of high
production of DHEA, Period A and C/D; these are also the times of the two
dentitions in humans. During growth of the "permanent" dentition,
front teeth develop during the final stages of brain growth. This competition
causes the front teeth to be small; as the process of brain growth finishes, the
size of the teeth increases. The very large molars develop during a time when
ample DHEA is present for growth. We lose our teeth during DHEA decline of
redundancy.
It is known that testosterone increases sex
drive in males and females. Since modern human females have sex throughout
their cycle while chimpanzees are limited to estrus, I suggest the difference
results from the increased T in modern women. Therefore, female hominids of
increased T had a selection advantage in reproduction. They increase the
probability of male attention throughout their cycles, and, therefore, increase
their reproduction rate. As the population of higher T female hominids
increased, their size would increase and the male-female difference would
decrease.
"Another correlate of brain
size is a decrease in male- female body size difference. Sexual dimorphism
remained marked in the pithecanthropines [now called Homo erectus], but
it is reduced from its Australopithecine extreme. The reduction in dimorphism
was not caused by a decrease in male size and robustness, but rather an
increase in female size." (The Stages of Human Evolution and Cultural
Origins, 3rd. ed., G.L. Brace, Prentice-Hall, 1968, page 93)
As testosterone increases in females, the
effect of estrogen declines in proportion. Estradiol in female humans and
chimpanzees is about equal, however, female chimpanzees announce sexual
receptivity with an extreme estrus display. Therefore, I suggest increases in T
in hominids reduced estrus displays, while, at the same time increasing sex
drive. Human female pubic and axillary hair is due primarily to adrenal
androgens, primarily DHEA. Since chimps produce more DHEA and hair than humans,
I suggest our relative lack of hair results from the T-target tissue
competition. That is, our hair is reduced because of reduced DHEA.
As testosterone increased in hominid females,
along with upright locomotion and reduced hair, competition among females must
have increased, especially with increased sex drive. I suggest this produced a
selection pressure for development of the breast as a primary sexual attractive
device; the same mechanism that produced the estrus display in chimpanzees. We
are the only group of mammals that use the breast as a sexual display. Breast
development is directly tied to the abundant form of DHEA, called DHEA sulfate,
from which DHEA is made.
"A significant positive
correlation was observed between DHA-S [DHEA sulfate], body weight and each
stage of breast development before and after onset of menarche." (Acta
Obstet. Gynaecol. Jpn. 1988; 40: 561)
The human breast display is directly related
to sexual maturity, i.e., ovarian function. The ovaries are connected to DHEAS
production: "Our data show that premature ovarian failure and ovariectomy
in young as well as postmenopausal subjects precipitate an earlier decline in
DS [DHEAS] levels" (Journal of Clinical Endocrinology and Metabolism
1982; 54: 1069.)
Conclusion
Human DNA and chimpanzee DNA differ by only
1.2%. This difference has taken six million years to produce. The DNA of
archaic Homo sapiens, H. erectus, and even Australopithecus
must have been even more similar to ours. Hominid evolution is a pattern change
more than a genetic change. I suggest it results from changes in hormone
production and their effects on gene regulation. Some genes have increased
activity, while others have decreased activity. These have produced significant
physical and behavioral changes over time.
Human evolution relies on simple changes in
hormone production, that result from basic behaviors that we see everyday.
Human evolution is viable and unyielding today, and affects every aspect of our
lives. In the next part of this series, I will explain how this mechanism
applies to contemporary society. This will be followed by a number of articles
concerning other hypotheses.
Further Support of
Theory
A number of newsgroup posts have connected
hair loss and sweat glands in the development of Homo sapiens. Often
these explanations deal with temperature. Since I think human evolution is
mainly the result of the increased testosterone in us, I must be able to show
that hair loss is due to increased testosterone and that sweat glands are a
target tissue for testosterone. We have less hair and more sweat glands.
If I am correct that we produce less hair
because of more testosterone, then reducing testosterone should increase the
amount of hair growth. This has been done in the stumptail macaque. In the
following quotation, note that "finasteride, a 5 alpha-reductase
inhibitor," significantly increases hair growth. Finasteride reduces the
effects of testosterone. That is, 5-alpha-reductase produces
5-alpha-dihydrotestosterone from testosterone in "testosterone target
tissues." If this enzyme product of testosterone is reduced, hair growth
increases.
"Finasteride, a 5
alpha-reductase inhibitor, was administered orally (1 mg/kg.day) for 6 months
to six male and five female stumptail macaques. Vehicle was given to five male
and five female animals over the same period of time. Hair weights in a defined
1-in.2 area of frontal scalp were measured periodically every 1-2 months, and
serum was collected for measurement of testosterone and dihydrotestosterone. In
addition, scalp biopsies were taken before and 6 months after treatment to
evaluate the micromorphometry of hair follicles. Results showed that both male
and female serum dihydrotestosterone levels were significantly reduced (60-70%)
by finasteride treatment. Both males and females showed statistically significant
increases in mean hair weight over the treatment period compared to controls (P
= 0.034). In addition, there was a statistically significant increase in mean
follicle length (measured histologically in scalp biopsies) compared to
baseline in the finasteride-treated animals (P = 0.028)." (J. Clin.
Endocrinol. Metab. 1994; 79: 991)
In the stumptail macaque, reducing the
effects of testosterone increases hair. So, increases in testosterone in Homo
sapiens may be the reason for reduced hair. Different areas of hair growth
respond to testosterone in differing amounts. "Androgens [testosterone]
stimulate hair growth in some areas, e.g., beard, but may cause regression and
baldness in the scalp" [Clin. Endocrinol. (Oxf.) 1993; 39:
633.] My basic principle, of my work, is that the hormone, DHEA, is used in
transcription and replication of genes. (DHEA is used to "read" genes
for gene activity and copy genes for equal distribution in cell division.) I
have suggested that tissues differ in their use of DHEA; this is how I explain
evolution of eukaryotes and multicellularity. Therefore, tissues will require
different levels of DHEA for specific gene expression. Scalp hair and beard
hair are examples of this. I suggest the differentiating factor is the availability
of DHEA. It has been found that the receptor for DHEA can bind
dihydrotestosterone (the 5 alpha-reductase product) secondarily. That is,
"Bound [3H]DHEA was displaced sensitively by DHEA and secondarily by
dihydrotestosterone, but not effectively by other steroids, including DHEA
sulfate" (J. Clin. Endocrinol. Metab. 1995; 80: 2993.) This
means, to me, that DHEA is absorbed for growth of hair primarily, but the
by-product of testosterone, dihydrotestosterone, can compete for its receptor.
(This should happen at the cell surface and within the cell.) Therefore,
expression of genes dependent on less DHEA will be adversely affected by the
presence of dihydrotestosterone. This is why increased testosterone reduces
hair over the body, but not the hair producing tissues of the face.
Hair is present from birth. Since DHEA is at
its highest immediately following birth, some neonates of high DHEA should have
hair at birth. However, since the brain, primarily, and body start to use so
much DHEA for growth and development (gene function and replication), the DHEA
falls quickly after birth and the original hair is lost. (See my chart of DHEA
during the human life-span above.) This is the same reason that the deciduous
teeth form early, then are lost.
I have explained, just above, that tissues
differ in their dependence on DHEA. Testosterone target tissues have their
testosterone target genes "turned on" by testosterone. These genes
then use DHEA for transcription. Following the finalization of brain growth,
DHEA begins to increase in amounts in the blood from late childhood (5-7
years); this is called adrenarche in the textbooks. (The textbooks do not have
an explanation for this.) What this means to this discussion is that DHEA
begins to increase from late childhood to reach a peak around 20 to twenty-five
years. Since sweat gland activity really begins following puberty, I think this
means that the rise in testosterone in men and women is the cause. Sweat glands
are a phenomenon of testosterone, and this is an affect on gene activity.
"1. To study the difference in
sweat rate between men and women the rates of cholinergic-induced sweating were
measured in normal people before and after puberty and in response to androgens
and anti-androgens. 2. Sweat rate in men was more than double that in women. 3.
This difference did not occur in prepubertal boys and girls in whom the rate,
corrected for surface area, was comparable with that in women. 4. Application
or injection of androgen locally did not stimulate sweat production in the
adult female. 5. Anti-androgen topically or systemically did not decrease sweat
rate in men. 6. It is concluded that the rate of sweat rate in men is caused by
androgen-induced gene expression at puberty and not by androgen modulation in
adult life." (Clin. Sci. 1981; 60: 689)
The next quotation demonstrates that sweat
glands have the highest 5 alpha-reductase activity of the entire skin,
sebaceous glands have a high activity, and hair follicles have significantly
less activity than the sebaceous glands. As you read this, think about the
increased activity in males, that may, therefore, increase the activity of the
sweat glands, which could further increase hair loss in the scalp.
"In order to know the
distribution of testosterone 5 alpha-reductase activity in human skin, we
developed a micro-method, in which we used 20-50 micrograms of various tissues
microdissected from freeze-dried sections. The characteristics of this enzyme
in the sebaceous gland are briefly described, as follows: the identified 5
alpha-reduced metabolites are 5 alpha-dihydrotestosterone, 5 alpha-androstane-3
beta, 17 beta-diol and 5 alpha-androstanedione; the optimal pH is about 7.5;
and the apparent Km is approximately 2.4 x 10(-5)M. The measurement of 5
alpha-reductase activity of various components of the skin obtained from 7 men
and 5 women revealed that the sweat gland (probably apocrine) in the axillary
skin possessed the highest activity of 5 alpha-reductase: the value was nearly
400 pmoles/mg dry weight/hr in the standardized condition. The sebaceous gland
also showed a high activity of 85-261 pmoles/mg/hr. The hair follicles
exhibited a significantly lower activity than the sebaceous gland. The enzyme
activity was negligible in the epidermis, while it was detected in the dermis
though the values determined were variable probably because of contamination
with other components such as sweat glands and hair follicles. Thus, the
present study demonstrates that the 5 alpha-reductase activity is mainly
located in the apocrine sweat gland and sebaceous gland. This suggests that 5
alpha-reduction of testosterone is an important step in mediating the action of
androgens in these tissues." (J. Invest. Dermatol. 1980; 74:
187)
Testosterone is known to increase sex drive
in both males and females. This would increase the percentage of higher
testosterone hominids with time. Increased testosterone would reduce hair,
increase sweat glands and activity and, in the female would reduce labial
displays, normally dependent upon increased estrogen to testosterone. The
exposed breast, also indicative of sexual maturity, would become the primary
sexual display. This combination would eventually lead to bipedalism. Other
events, dependent upon the hormones DHEA and melatonin, would, much later, result
in an enlarged brain.
So, you see, one does not have to resort to
looking for environmental effects to account for all of these characteristics
of hominids. The single mechanism of increases in testosterone, alone, will
cause all of these changes. That is, increases in testosterone increase the
sexual device. The sexual device is one of most important devices created by
DNA for duplication.
Current Signs of
Increases in Testosterone in the U.S.
Testosterone is the basis of violent
behavior. That is, testosterone is the basis of impulsive behavior. The amount
of testosterone determines the ability to control, or not, impulses. More men
are imprisoned than women. Black men (at the college level) produce more
testosterone than white men; more black men are imprisoned than white men. The
following is a letter describing this, which has been sent to a number of U.S.
congressmen and U.S. senators. You judge for yourself. This is from 1994.
"I am a theoretical biologist;
my work contains an explanation of increased violence in our society. I suggest
violence results directly from an increase in numbers of individuals of higher
testosterone, who arrive at puberty early. increased testosterone and early
puberty increase the probability of impulsive actions. The Federal Bureau of
Investigation has compiled statistics which demonstrate that I am correct.
Males kill more than females; blacks kill more than whites. In the remainder of
this letter, I include references from highly reputable journals (e.g., Journal
of the National Cancer Institute) that demonstrate the blacks produce
significantly more testosterone than whites and enter puberty at an earlier
age. What this boils down to is that individuals of both races, who exhibit
these qualities, are more apt to resort to violence in a heated moment. This is
why there is so much black on black and white on white violence; these are
impulsive actions, not premeditated, thoughtful actions. These are thoughtless
actions that happen quickly, without forethought. You, or your aides, are aware
from interviews that many of these kids, who kill other kids, are really nice
kids when they are not in a stressful, heated moment.
I am aware your background is not
biological, nor are the backgrounds of those whose advice you seek when
confronting the increase in violence in our society. You must consider,
therefore, that a major explanation of human behavior is being neglected by
educators, sociologists, criminologists, etc. Their "model" of human
behavior suggests that almost all human behavior is determined by the
environment. It is time a biological model is considered. I will explain how
this produces violence in the remainder of this document; if you read this, you
should note the same mechanism also explains increased sexuality and learning
problems in our youth. That is, all of these result from a single change in our
society, which is very easily produced and increases exponentially.
This change increases with each
generation. Not everyone is affected, but more children are affected now than
in the past. Therefore, each past generation contains more who notice the
change. More grandparents see it than parents. It occurs earlier in some places
than others. This is why some people always see these problems in some place at
some earlier time. I think the "60s" was the time it occurred in such
magnitudes that it openly impacted our entire society. It is a continuing
process, but usually takes more time to occur in conservative areas.
My works suggests this is a major
biological change that affects both the body and brain. Most grandparents and
some parents have noticed the change in body size and function in children.
Children are getting bigger and reaching puberty earlier. "The average age
of menarche [puberty] in the female has dropped from approximately 17 years to
approximately 13 years. Thus, today maturation occurs about 25% faster than it
did 100 years age" (Sexual Dimorphism in Homo sapiens, R.L. Hall,
Praeger Publishers, New York, 1982, page 279.)
"...children in average
economic circumstances have increased in height at age five to seven by about 1
to 2 centimeters per decade. ...Most of the trend toward greater size in
children reflects a more rapid maturation; only a minor part reflects a greater
ultimate size." (Encyclopædia Britannica 1984; 5: 656)
My work suggests a cause of this
change. The hormone, testosterone, is rising rapidly in our society. Increased
testosterone increases body size, aggression, and sexuality in both sexes.
(Testosterone is not "the" male hormone, men simply produce more.)
People who produce more testosterone are more aggressive and sexual, therefore,
on average, they ultimately make more babies than those who produce less
testosterone. (People who produce less testosterone can better control their
sexual activity; over a period of time, they will produce fewer children.)
Ultimately, the percentage of high testosterone people, of both sexes,
increases at the expense of low testosterone people. This changes the averages of
everything affected by testosterone. This is why our kids are bigger, more
sexual, and more aggressive than in the past.
The mechanism is simple: higher
testosterone boys and girls reach sexual maturity faster, increase their
numbers faster, and their offspring are even earlier and more sexual. People
seeking sexual gratification are simply more likely to engage each other.
Sexual activity is so common today that no "stigma" is attached; in
fact, there appears to be a negative stigma attached to those who do not
indulge.
Prior to puberty, the brain grows
more rapidly than the body; it is a competition which the brain wins in infancy
and early childhood. Because of this brain-body competition, puberty is delayed
until the brain is almost finished in development. Near puberty, however,
testosterone increases the body's competitive edge for growth and development
which continues into adulthood. "The weight of the brain [in humans]
reaches 90% of adult size by age six and virtually 100% by age 12, yet body growth
continues to age 18 and beyond (note that brain growth is nearly finished
before reproductive maturity every begins)" (Patterns of Human Growth,
Cambridge University Press, 1988, pages 60-61.)
The advanced frontal lobes of the
brain develop last and control formal thinking, i.e., higher math, proper
language (syntax), and the ability to form meaningfully predictive ideas
(hypotheses). This is Piaget's final stage of human thought development. This
stage of brain development is directly dependent on final development of the
frontal lobes
"from
about age 11 to 14," (Science 1987; 236: 1110). I suggest
early puberty interferes with this important final development of the frontal
lobes. For example, it was reported that standardized test scores of 13- and 17-year-olds
of 1986 are lower than those of 1970, whereas the scores of 9-year-old children
have remained relatively equal (Science 1988; 241:1751). I
suggest this decline is the effect of puberty, which, in this country, on
average, is now occurring between age 9 and 13. Our children are, on average,
losing the ability to handle math and English. More importantly, our children
are losing the ability to form meaningfully predictive ideas that help control
their impulses. "What are the consequences of my actions?" Without
the function of the frontal lobes, symbolized by this question, kids cannot
predict the consequences of, or control, their behaviors (impulses). Violent
acts and sexual activity in children and teenagers are actions of impulse.
These impulses are initiated by the primitive part of our brains, which
testosterone mainly affects. Children are reaching puberty earlier with each
generation, and early puberty arrests final development of the brain. This
means that, on average, our advanced brain is increasingly underdeveloped with
each generation. This is why so many children cannot control their sexual or
aggressive impulses.
It is fact that, on average, the
behaviors mentioned above, occur in higher incidence in the black population.
That is, on average, black children have more problems with math and English,
score lower on standardized tests, exhibit more aggressive and sexual impulse
activity, and experience family disintegration more than white children. The
reason, I suggest, is that, on average, blacks, as a group, produce more
testosterone than whites, as a group. (Take note of the journal source of the
following quotation.)
"This report gives the results
of assays of circulating steroid hormone levels in white and black college
students in Los Angeles, CA. Mean testosterone levels in blacks were 19% higher
than whites, and free testosterone levels were 21% higher. Both these
differences were statistically significant. Adjustment by analysis of
covariance for time of sampling, age, weight, alcohol use, cigarette smoking,
and use of prescription drugs somewhat reduced the differences. After these
adjustments were made, blacks had a 15% higher testosterone level and a 13%
higher free testosterone level." (Journal of the National Cancer
Institute 1986; 76: 45)
The study, above, was of college
students. If I am correct that testosterone adversely affects learning, then
those blacks who are unable to meet requirements for college admission, or even
complete high school, may produce more testosterone, on average. The should
exhibit even less impulse control; this may be why there are so many black on
black murders in ghettos. This was not a common occurrence in black communities
in the depression. Testosterone is increasing over time.
I have suggested that increases in
testosterone in our society are causing the overall problems, i.e., increases
in testosterone are causing problems for both blacks and whites. Therefore, the
rate of teenage births should be higher in blacks than whites but increased in
both, compared to other advanced countries. This is the case.
"The rate of teenage births is
especially high in the black population. An international comparison around
1980 revealed that the black U.S. teenage fertility rate was 2.3 times the
white and 3.2 times the average of 30 advanced countries. The contrast is even
greater among the youngest teenagers; blacks under 18 years of age had a rate
in 1980 that was more than three times that of whites. Nonetheless, even white
teenage birthrates were 40% higher than the average for other advanced
countries." (Science 1986; 234: 554)
Not only do blacks, as a group,
produce more testosterone, which I have suggested, increases the onset of
puberty, they do, in fact, reach puberty earlier than whites as a group.
"Black youths are known to
enter into puberty at a younger chronological age than white youth." (American
Journal of Diseases of Children 1991; 145: 142)
Again, it is my hypothesis that the
violence, sexuality, and learning problems of our youth result from increased
testosterone and early puberty in those affected. High testosterone and early
puberty adversely affects development of the part of the brain which controls
impulsive behaviors, i.e., the advanced forebrain. This combination should
generate these problems, and they should be exacerbated in areas where high
sexuality rapidly brings high testosterone males and females together. The
result is an extremely rapid increase in high testosterone, early puberty, and
their combined effects on impulse control. As people of lower testosterone are
literally driven away, the problem becomes more concentrated. Impulsive acts
also become concentrated; this is why there is so much black on black and white
on white violence."
A Letter to the
Editor
Northwest Arkansas Times, Fayetteville, AR, U.S.A., May 22, 1998.
"May 18, page A3, the ... [Northwest
Arkansas] Times... reported Retired Army Lt. Col. Dave Grossman's
conclusions that "Like military training that reduced inhibitions to
killing, television and movie violence is desensitizing the young, and doubling
the murder rate every 15 years." This is not a new idea; many people have
suggested the "media" causes increased violence and sex in our youth.
It produces a "straw man," with which one may do lengthy, imaginary
battle. My work suggests the current phenomena, and others, are the result of
human evolution. That is, I think humans evolved due to increased testosterone.
Human males and females produce more testosterone than male and female
chimpanzees, respectively. In advantageous circumstances, testosterone levels
will increase in a population. Increased testosterone increases impulsive
behavior. We are witnessing an increase in testosterone in America.
This is directly supported by very strong
correlative data and direct experimental evidence. In a study of
"delinquent" and "control" white men and women, Banks and
Dabbs found that "The delinquent group, which was characterized by
flamboyant dress, drug use, and violence, had significantly higher testosterone
levels than the college students did." (J. Soc. Psychol. 1996; 136:
49). Brooks and Reddon compared testosterone levels in violent and nonviolent
"young offenders." They found that "The violent group had the
highest level of testosterone and differed significantly from the nonviolent
offenders..." (J. Clin. Psychol. 1996; 52: 475). This
represents a strong correlation between high testosterone and impulsive,
aggressive acts. People who do not think aggressive, impulsive acts are due to
increased testosterone can simply dismiss this as coincidence.
The key experiment that directly supports
testosterone as the causative agent was reported in 1997. In a study of
"hypogonadal," boys, who produce little testosterone, the effects of
increased testosterone become clear. Finkelstein, et al., found that "At
the mid dose boys showed a 19% increase in aggressive impulses scores, a 17%
increase in physical aggression against peers score, and an 18% increase in
physical aggression against adults scores." (J. Clin. Endocrinol.
Metab. 1997; 82: 2433). Men produce much more testosterone than
women. There are many more men in prison than women.
My work suggests testosterone increases
periodically in civilizations. That is, where food and shelter are beneficial,
people of higher testosterone will increase rapidly, compared to low
testosterone people. They are more sexual and impulsive; they make more babies.
They are bigger and reach puberty earlier; this is known as the secular trend.
The secular trend is not due to better food. Black girls reach puberty much
earlier than white girls, and there is no support that black girls eat better
than white girls. Impulsive acts will increase directly proportional to the
increased numbers of higher testosterone types. High testosterone should have a
greater effect on a kid following puberty, because the brain is not fully
developed. The greatest amount of youth violence is coming from young, male
offenders. Let's look at biological factors."